
Metabolism and Metabolic markers of Dementia and Alzheimer's Disease
The majority of metabolic markers that are common in dementia and Alzheimer's disease, are also associated with neuronal degradation, and also associated with absolute or functional vitamin B12 deficiency. Careful monitoring of these markers potentially could result in prevention of dementia.
-
Elevated homocysteine- an associative marker for functional vitamin B12 deficiency
-
Decreased creatine - a methylation product, which is reduced in functional B12 deficiency
-
Decreased GSH:GSSG ratio => Protein misfolding
-
Decreased production of hydrogen-sulphide - and essential neurotransmitter, a product of the sulphation cycle
-
Sleep disorders - due to reduced production of the methylation product, melatonin
-
Reduced activity of the iron-sulphur protein aconitase - an indicator of iron and B12 deficiency
-
Reduced myelination and repair- due to lack of melatonin and vitamin D deficiency.
-
Reduced levels of vitamin D - Vitamin D works in combination with melatonin in the differentiation of neuronal stem cells to myelin-producing oligodendrocytes
-
Frailty - due to iron and functional B12 deficiency
Apart from the overt "mental" signs of early Alzheimer's disease (AD), it has now been shown that there are definitive metabolic signs of AD. Analysis of the serum and urine of persons with dementia and Alzheimer's disease has shown some significant differences between normal healthy individuals and those with developing cognitive impairment and Alzheimer's disease. Whilst these markers may not necessarily be definitive for diagnosis of Alzheimer's disease it should be possible to plot the progression or initiation of the disease by measurement or the markers, and knowledge of what the markers "mean" may suggest possible causes for the development of the symptomatology of AD. Some of these markers are outlined below, with an explanation of what they mean or how they correlate with the condition. The explanations are quite detailed however, they do show that it should be possible to measure or detect changes in metabolism along the "journey", by using the markers.

Figure. Increase in Homocysteine with Age (Seshadri etal, (2)
Homocysteine: Homocysteine is an unusual amino acid, not coded for by DNA, and not found in proteins, but rather it is produced as a metabolic by-product of the methylation cycle. In normal individuals homocysteine is either processed to form methionine by the methyl-vitamin B12 dependent enzyme, methionine, synthase, or it is taken into the sulphation cycle by processing of an enzyme called cystathionine beta synthase (CBS). Elevated homocysteine has been shown to be an indirect risk factor for vascular disease and for dementia in older people (1,2). It is also associated with hypertension (3). Alzheimer's disease progression is associated with increasing hyper-homocysteinemia (elevated homocysteine) and vascular dementia (4,5). It has also been associated with thinner cortical gray matter (6) decreased hippocampal volume (7) and brain atrophy (8). Elevated homocysteine has also been associated with reduced production of the vasodilators nitric oxide and hydrogen sulfide (H2S) (9). Elevated homocysteine has also been associated with a decrease in Mini Mental Score Estimation (MMSE) values in AD (10-14). Typically the elevated homocysteine, per se, has been thought to be causally related to the above conditions, however, it is much more likely that the nutritional deficiencies in iron, folate, B12, vitamin B6, or functional vitamin B2 are the true causes of the conditions, and homocysteine is but a marker of such deficiencies. Studies have shown that homocysteine levels start to rise as serum ferritin levels fall below 50 ug/L.

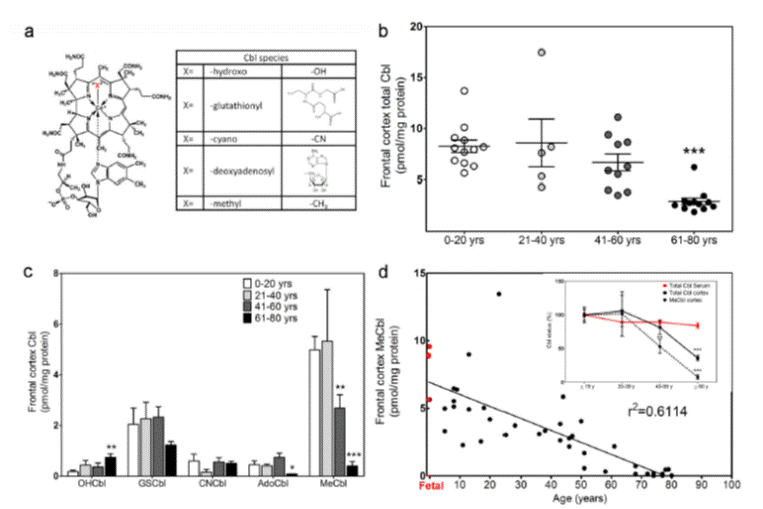
Figure. Decrease in methyl B12 in the frontal lobe of the brain during ageing (Data from Zhang etal, (18))
Reduced levels of vitamin B12 have also been associated with uncoupling of important iron-sulphur proteins, such as aconitase, succinate dehydrogenase, and complex III, in the important Krebs cycle and the electron transport chain. There is also a reduction in the activity of GABA aminotransferase, which may explain the depression, which often accompanies vitamin B12 deficiency. More recently, iron-sulphur proteins have also been implicated in the final activation step of vitamin D, in which 25-hydroxyvitamin D (calcidiol) is converted to the active molecule 1,25 dihydroxyvitamin D (calcitriol), through the interaction of CYP27A1, ferredoxin (an iron-sulphur protein) and ferredoxin reductase..
Vitamin B12: Vitamin B12 deficiency has been shown to be associated with frontotemporal dementia and increased memory loss (16). Levels of serum vitamin B12 below 300 pmol/L have been shown to result in increasing reduction in mental capacity as can be determined by Mini Mental Score Estimations (MMSE)(17). Vitamin B12 is essential for the formation of the myelin sheath around nerves and vitamin B12 deficiency has also been associated with myelin degeneration (18) . Levels of methyl B12 have been shown to be greatly reduced in the elderly brain (18) and significant improvement in cognitive function has been shown with intravenous administration with methyl B12 (19,20). Lack of myelination of nerves greatly reduces the speed at which they can carry information. Unfortunately, vitamin B12 deficiency is very commonly missed, possibly due to the reliance on the old haematological assessment of vitamin b12 deficiency, which was based on the early belief that the only role for vitamin B12 in the body is for the production of red blood cells, which technically it doesn't do. This method of assessment disregards the important role that vitamin B12 plays in maintaining methylation and in energy production, as such they will not treat patients for vitamin B12 deficiency unless they are below 150 pmol/L (21). This means that the many people who have early signs of vitamin B12 deficiency are not being treated for the condition, rather are treated with numerous drug cocktails for a variety of conditions, such as cardiomyopathy, depression, psychosis, impaired cognition, which if properly diagnosed at early onset would potentially halt the progression of dementia.
Reduced vitamin B12 levels impact upon energy production in mitochondria, and through it's essential role in methylation and the production of S-Adenosylmethionine, reduced methyl B12 also results in lower levels of creatine and creatine phosphate, and reduced activity of the electron transport chain due to lower production of ubiquinol (CoQ10). Reduced cognitive function is associated with lower levels of ubiquinone in several conditions including autism, Huntington's disease and dementia (22,23). Ubiquinol administration has been claimed to be neuroprotective in neurodegenerative diseases (23). Serum coenzyme Q10 levels have been touted as being a useful predictor of dementia (24,25,26). Over 40% of all methylation goes to the production of creatine. Low levels of creatine are a feature of AD. Low levels of acetylcholine in the brain.
Other B12 deficiency markers include HVA, VMA, QA, KA, 5HIAA, MMA, Pyroglutamate, reduced SAM:SAH ratio,
Further complicating the association of vitamin B12 with dementia, is the association between metformin use and vitamin B12 deficiency in patients with type 2 diabetes, where it has been found that on average there is a 57 pmol reduction in B12 levels after 6 weeks to 3 months of use (27).
In addition, it has been found that there is reduction in vitamin B12 levels as body mass index increases. This may be as a result of lack of functional B2 (as FMN and FAD), which would both reduce the ability to burn fat and also reduce B12 cycling.
Homocysteine: Functional vitamin B12 deficiency is associated with elevated homocysteine, as seen in AD, ASD and PD. This in turn is associated with increased cerebrospinal levels of Beta Amyloid protein. Supporting the concept of increased mis-folding of proteins in functional B12 deficiency.
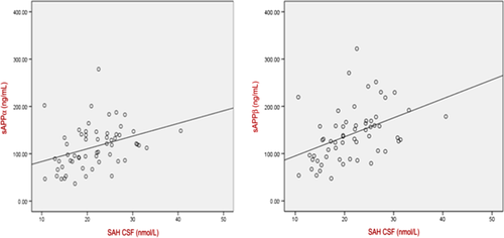
Oikonomidi et al, 2016
Creatine: Movement of ATP formed within the mitochondria, out into the cytoplasm of the cell is greatly enhanced by the action of membrane-bound creatine-kinase, using creatine as backbone to accept the high energy phosphate from ATP. Formation of creatine is dependent upon methylation of guanidinoacetate by the enzyme GNMT, which uses SAM as the methyl-donor. Over 40% of all methylation in the body is involved in the production of creatine. Executive function in geriatric depression was found to be correlated with levels of phosphocreatine (28). Creatine deficiency has been demonstrated in conditions such as Chronic Fatigue Syndrome, in which brain fog, is a common symptom, and in autism, in which the children have difficulties in speech, continence, and have sleep disorders, which are all conditions also associated with AD. Creatine deficiency is also a feature of Alzheimer's disease (Roschel etal, 2021; Osaki etal, 2022; Snow etal, 2020; Mihara etal, 2006).Creatine has also been shown to have an essential role in production of energy for myelination and remyelination of nerves by oligodendrocytes.
Creatine and energy production within the cell
The penultimate step in energy production within the cell is the transfer of ATP across the mitochondrial membrane via the enzyme creatine-kinase, to an awaiting creatine molecule in the cytoplasm of the cell to make the high energy phosphate donor Creatine-Phosphate. Without this step, the generation of ATP within the mitochondria is futile. Thus, the creatine/phosphocreatine shuttle system is an essential component of transport of energy, produced in the mitochondria, into the cytoplasm of the cell (Sacks et al, 1978) . As such, it is thought to be essential for storing of high phosphate-bound energy, particularly in those cells with high energy demand. Creatine levels are high in tissues such as muscles, the brain, and are also very high in the oligodendrocytes Braissant etal, 2007; 2008; 2011) and astrocytes. It has been known for some time that Creatine-kinase mRNA levels are high in oligodendrocytes and astrocytes Molloy etal, 1992. The methylating enzyme GAMT, which is involved in the final step in creatine synthesis is similarly found in these cells Tachikawa etal, 2004). Whist originally it was thought that most of the Creatine in the brain was of peripheral origin, more recently evidence suggests that the ability of creatine to cross the blood brain barrier is very poor, and hence the majority of Creatine used in the brain comes from endogenous synthesis (Braissant etal, 2007; 2008; 2011). This, then, potentially creates a problem in functional vitamin B12 deficiency, because the synthesis of Creatine in the brain will also require an active methylation cycle locally in the brain, to provide the methyl donor SAM for use by GAMT in the synthesis of Creatine.The corollary to this is that in functional B12 deficiency (shown above), there will be a reduced production of creatine, with resultant brain fog and difficulty in thinking, characteristic of AD.
Creatine supplementation and Improvement in Cognition
Studies on Vegan subjects given 5 gm per day creatine-monohydrate, showed a significant improvement in cognitive scores after 4 weeks of supplementation. The mechanism was presumed to be greater uptake of creatine into the brain and neuronal cells (Rae etal, 2003). A similar improvement was seen cognition by Hammet and co-workers (2010).
Iron: Depletion of serum ferritin (an iron carrying/storage protein) and iron accumulation in the brain has been found in AD (29-33). The level of iron deficiency as measured by serum iron and transferrin saturation correlated with lowering of cognitive scores (34). Iron accumulation in the brain has been posed as an initiating factor in the aggregation of beta amyloid, such as is seen in the brains of persons with advanced dementia (35). Iron dysregulation has also been proposed as a mechanism for the disruption of iron-sulfur biogenesis (35). Ferritin levels in the cerebrospinal fluid have been found to be predictors of AD disease outcomes (36), with increasing levels of ferritin in the brain being negatively associated with cognition. Diagnosing iron deficiency is very controversial and there are two distinct "camps" defining iron deficiency. One "camp", appears to believe that iron is only used for the production of the heme structure in proteins such as Haemoglobin. This group defines iron deficiency at the level when haem production is reduced. They thus define iron deficiency at 15-20 ug/L ferritin. This is despite evidence that even for heme-dependent enzymes, such as CBS and aconitase, activity starts to fall as ferritin values drop below 70 ug/L. It appears that these people are still using old reference values or text books possibly written by early physicians such as Hippocrates or Pedanius Dioscorides. These individuals, and in fact most of the medical text books and all the psychology text books that we could find, have totally ignored the non-haem uses for iron, viz, the formation of iron-sulphur complexes and the resultant iron-sulphur proteins, and the use of "singlet" iron in enzymes such as tyrosine hydroxylase. Formation of iron-sulphur complexes and the activity of iron-sulphur proteins is reduced a much higher concentrations of serum ferritin, and our data suggests that the inflection point for this (as judged by aconitase activity - a major enzyme in the Krebs cycle - see below) is at around 100 ug/L ferritin, which is the definition of many research groups to define iron deficiency (the second "camp). This is particularly important as recovery from heart failure improves as serum ferritin rises from 20 to 100 ug/L. In addition, there is a decrease in muscle iron as ferritin drops from 100 ug/L, potentially explaining the decrease in muscle strength seen in dementia. Further, as early as 1976, it was shown that muscle loss of iron preceded the development of anemia in rats (37). Iron deficiency in the brain precedes the signs of iron deficiency in RBC production. Serum ferritin levels below 76 ug/L are associated with abnormalities in neonatal recognition memory, and neuronal processing (38-44). Further, a recent study has shown that lower iron levels (as judged by serum ferritin) are associated with decreased brain activity and lower energy expenditure, as well as a reduced heart rate (45).Reduced iron stores in bone marrow are apparent when ferritin levels drop below 75 ug/L (46).Given that iron deficiency is associated with depression (a common symptom in dementia), which is possibly due to dissociation of the iron-histidine chelate within tryosine hydoxylase, it seems incongruous that the potential exists for health professionals to treat depression with anti-depressants who sit in "the first camp", whilst in all possibilities iron supplementation would be a much more successful/useful approach, particularly for those who have ferritin between 20-100 ug/L. This, though, might be at odds with the massive US$14 billion spent on anti-depressants in 2014. Perhaps it is time for pathology labs to move on from defining iron deficiency in terms of oxygen carrying capacity of red blood cells and move into the real world where iron is essential for energy metabolism, and synthesis of neurotransmitters such as dopamine and serotonin, and so adjust their ranges to reflect more accurately the true values for iron deficiency. Interestingly, in discussions that we have with several labs working on iron deficiency, they have deliberately set their definition of iron deficiency to less than 15 ug/L ferritin, as they were worried that if they lifted their cut-off values higher, too many of their subjects would be deemed iron deficient
.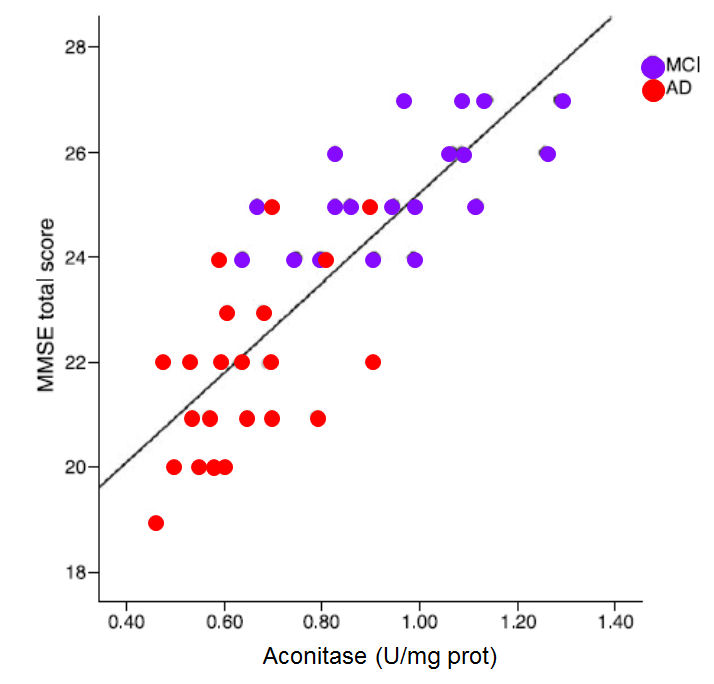
In the graph the MMSE score is plotted against the activity of the enzyme aconitase (Figure. Data from Mangialasche etal, (39)
Sleep Disorders.: Sleep disorders are associated with cognitive decline and are common to neurodegenerative diseases such as dementia and Alzheimer's disease Holingue etal, 2018; Mattis and Sehgal, 2016; Blackman et al, 2020; Porter etal, 2015; Chen etal, 2015; Henry etal, 2019; Sabia etal, 2021; Peter-Derex etal, 2015). One of the most common symptoms of vitamin B12 deficiency is sleep disorders. The main sleep hormone is the methylation product, melatonin, which is formed by methylation of N-Acetyl-serotonin. Children with sleep disorders as a child are often B12 deficient. Of particular relevance to dementia, is the observation that m Melatonin, in combination with vitamin D is intrinsically involved in the activation of neuronal stem cells, into myelin-producing oligodendrocytes (see below).."Endogenous melatonin levels are inversely associated with age and cognitive defects" (Sumsazzman etal, 2021).
Aconitase: Aconitase is one of the first enzymes in the citric acid cycle (Krebs cycle) that is involved in processing acetyl-CoA, the common metabolic break-down product of fats, sugars and proteins. Decreased activity of aconitase has been found to parallel the drop in Mini Mental Score in progressive dementia and Alzheimer's disease (46-48). Reduced aconitase activity has been shown to correlate with ageing (49). Reduced usage of acetyl-CoA by the CAC in AD is apparent by the finding of increased levels of Acetyl-CoA in the hippocampal cortex in AD patients when compared to controls (50). Reduced consumption of acetyl-CoA for energy also leads to an increase in cholesterol, which is also common in AD (14). Reduced aconitase activity can be measured by increasing levels of citric acid in the urine.
Aconitase activity is also a surrogate marker for the integrity of iron-sulphur proteins in general, hence as aconitase activity drops so too will the level of activity of GABA-aminotransferase, Ferredoxin, Lipoate Synthase, and Xanthine oxidase.
Acetylcholine: Acetylcholine is a major neurotransmitter involved in short term memory and in transmission of information from nerves to muscles. AD is associated with the loss of cholinergic neurons (nerves that respond to acetyl choline) in the brain, which may result from the decreased levels of acetylcholine in the brains of the AD individual. Increasing the level and persistence of acetylcholine in the brain is one of the major therapies used in treatment of AD. Acetylcholine is normally broken down by acetylcholine esterase (AChE) and AChE inhibitors have been approved for use in the treatment of AD, albeit with little success (50). Unfortunately, to date treatment has not addressed the basic question of why the levels are reduced. One of the best sources of choline are eggs, and there is a reduced risk of Alzheimer's disease in those with a higher egg intake (Pan et al, 2024).
GSH:GSSG ratio: Glutathione is essential for reduction of metals such as iron, and copper, and reduced levels of free GSH are found in AD. The GSH:GSSG ratio within the cell also determines the extent of "disulfide shuffling" seen during the correct folding of proteins produced within the cells. Protein folding is the process by which a polypeptide chain acquires its functional, native 3D structure. Protein misfolding, on the other hand, is a process in which protein fails to fold into its native functional conformation. This mis-folding of proteins may lead to precipitation of a number of serious diseases such as Cystic Fibrosis (CF), Alzheimer's Disease (AD), Parkinson's Disease (PD), and Amyotrophic Lateral Sclerosis (ALS), Autism Spectrum Disorder etc. (Yadav et al, 2019; Gamez et al, 2018, Wankhede et al, 2022) This is postulated to be one of the main reasons for the occurrence of protein aggregates in the brain, such as those containing Tau and Beta Amyloid. Alterred GSH:GSSG ratio correlates with iron precipitation in the brains. The reduced ratio of GSH:GSSG is a result of lower levels of active vitamin B2, as FAD, which is used as a co-factor by glutathione reductase.
Hydrogen sulfide: Hydrogen sulfide (H2S) is an important gaseous neurotransmitters produced in the brain. Production of H2S has been shown to be reduced in AD (53). Several studies have shown a potential therapeutic use of H2S in improving memory in AD (53-55) and in reducing Aβ amyloid plaques (50). Reduced production of H2S has also been found in hyperhomocysteinemia (9,48). Production of H2S is dependent upon movement of dietary sulphur resident in methionine, through the enzyme CBS and into the sulphation cycle. Such movement is reduced in B12 deficiency.
Cytochrome C Oxidase. Cytochrome C oxidase catalyzes the last step in the mitochondrial electron transfer chain, in which the energy generated by the electron transport chain is converted to ATP. The activity of this enzyme has been shown to be reduced in AD (51). Such a drop in activity is correlated with reduced energy production. The enzyme has heme iron and copper as co-factors, and reduced activity would be expected in a deficiency of any of these co-factors.
Bioenergetics. Factors such as lack of Cytochrome-C oxidase and aconitase function, iron deficiency and vitamin B12 deficiency would all compound to greatly reduce the bioenergetic output common to dementia, and particularly late onset Alzheimer's disease (LOAD), as recently observed by Cohen and co-workers (52). Thus, energy input into the Krebs cycle becomes increasingly small due to lack of aconitase activity, energy transfer out of Krebs into the Electron Transport chain (ETC) would be reduced due to lack of activity of succinate dehydrogenase, and electron transfer within the ETC would slowly grind to a halt due to lack of CoQ10. What ATP that is produced would be compromised by the lack of production of creatine. Effectively this would mean that the entire brain would gradually become starved of energy. Lack of creatine production, alone, has been associated with decreased mental function, such as is seen in cretinism. It is hard to imagine why one would need to explore further to understand the mental decay in AD.
Melatonin: Melatonin levels in the body are highest immediately after birth and for the first 5 years of life, after that they drop off with time, until by age 60 the levels are less than one tenth those seen at 3 years old. Potentially this, alone, can explain the deterioration in the brain as we age. Hence, it would almost be like an apoptotic event, or programmed cell death. Potentially, this, when combined with the lack of vitamin B12 in the brain could explain the inevitable progression of Alzheimer's disease.
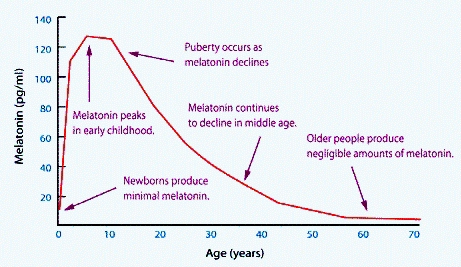
Melatonin levels during development (Grivas and Savvidou, 2007)
Nitric oxide: Nitric oxide is synthesized by the enzyme nitric oxide synthase (NOS). Vascular blood flow can be increased by the activity of endothelial nitric oxide synthase (eNOS), which synthesizes the powerful vasodilator, nitric oxide (NO). Reduced cerebral blood flow is a feature of AD, which would in turn reduce the flow of nutrients to the brain as well as restrict the removal of metabolic by-products from the brain. Studies by Selley (57) have shown that there is a highly significant decrease in the plasma concentrations of NO in AD, which is negatively correlated with Hcy concentrations. Reduced NO production has also been correlated with elevated homocysteine and hypertensive disorders (56), both are features of dementia.
Serotonin: Reduced levels of serotonin have been found in the serum of persons with AD (59-63). This may be reflected in the increased use of anti-depressants in persons with AD (54).
Selenium. Selenium is an essential co-factor (enzyme "helper") for the a number of enzymes, including Glutathione Peroxidase and formate dehydrogenase, but more importantly it is involved in the deiodination of the thyroid hormone T4 to form T3. T3 in turn is involved in the induction of riboflavin kinase, to convert riboflavin (vitamin B2) to one of the two active forms of the vitamin, FMN, whilst molybdenum is required for the formation of the second active form of the vitamin, FAD. Without these reactions, dietary or supplementation vitamin B2 is almost useless and functional B2 deficiency results. Studies by Cardoso and others (65,66) have shown that the selenium status in the elderly is related to cognitive decline, with erythrocyte selenium (red blood cell Se) significantly lower in elderly with AD, than in those in the control group. Persons with mild cognitive impairment have reduced Se when compared to controls, yet higher than the AD group. Studies in mice have shown increased production of amyloid-B plaques in selenium-deficient mice (67)
Demyelination. Myelin loss is a consistently seen in AD, both in the central nervous system and in the peripheral nerves, particularly in those nerves involved in proprioception (68). Lack of iron, vitamin D and vitamin B12 potentially contribute to the inability of oligodendrocytes to maintain and repair the myelin sheath. Evidence suggests (68) that white matter hyperintensities, characteristic of AD, are associated with myelin loss, and the loss of myelinated axons. White matter in the brain of AD patients reveals decreased amounts of total protein, myelin basic protein (MBP) myelin proteolipid protein and cholesterol. Differentiation of oligodendrocyte precursor cells is dependent upon the action of melatonin, a neuropeptide hormone whose levels are decreased in vitamin B12 deficiency. Neuronal circuits with higher neuronal activity are "given preference" in myelination, supporting the "use it or lose it" concept of AD. Apart from the need for melatonin, oligodendrocytes require 2-3 times the energy level of other cells in the CNS and are highly dependent upon iron for energy production in Krebs cycle and the CNS. Depletion in glutathione reductase activity has the effect of reducing available Fe++ for production of iron-sulphur proteins in the CNS.It is known that melatonin levels are lower in AD (Xu etal, 2015; Sumsazzman etal, 2021; Tseng etal, 2022), and "Endogenous melatonin levels are inversely associated with age and cognitive defects" (Sumsazzman etal, 2021). Melatonin has also been shown to have neuroregenerative properties (Potes etal, 2023).
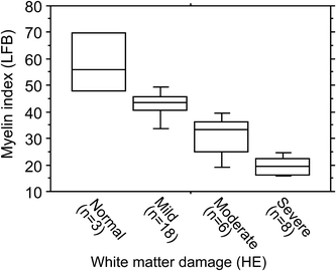
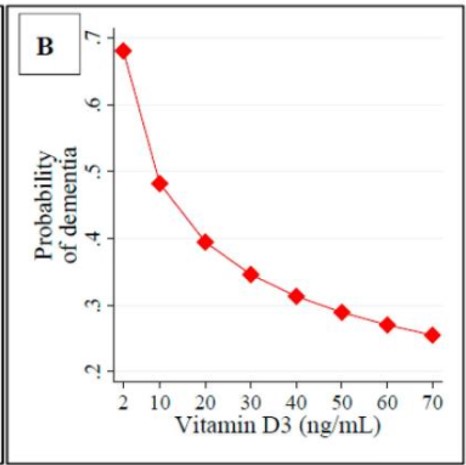
Vitamin D and Dementia. As described above, vitamin D is essential for neuronal differentiation and for myelination of nerves. In vitamin D deficiency at birth myelination is delayed. Vitamin D has also shown promise in neuroregeneration in models of AD (Morella etal, 2018;; Nasrabady etal, 2015; Gáll etal, 2021); Landel etal, 2015; Szczechowiak etal, 2015) Vitamin D deficiency has been associated with neurodegenerative diseases such as dementia, PD, and AD (Gail, etal, 2021). Under the Nexus theory of vitamin D activation, all of iron, B2, B12, and vitamin D are required for maintenance of neuronal function, and lack of any of these will result in gradual neuronal degradation and eventual loss of function.
Vascular disease, hypertension and Dementia. Hypertension (high blood pressure) is prevalent risk factor for stroke (blockage of blood flow, or rupture of an artery to or in the brain), and may contribute to the development of dementia due to lack of oxygen at the site of the event, or subsequent neuroinflammation (69). Several studies have shown that metabolic vitamin B12 deficiency and elevated homocysteine is common in Large artery artheroscleerosis ,small vessel disease and cardioembolic atherosclerosis (70), suggesting an as yet unidentified role for vitamin B12 in these conditions.
Glucose hypometabolism. Metabolism of glucose ultimately finishes at the enzyme pyruvate decarboxylase, a vitamin B1, B2 and lipoate dependent enzyme. Deficiency of vitamin B2 leads to hypometabolism of glucose (as is seen in diabetes) and build up of lactic acid. Studies looking at cerebrospinal fluid of patients with Alzheimer's disease have shown hypometabolism of glucose and increased production of lactic acid, commensurate with a block in activity of the enzyme, similar to that observed in vitamin B2 deficiency (71, 72). It is well known that the decreased brain utilization of glucose always accompanies the development of AD and precedes clinical manifestations by many years (73, 74)
Mitochondrial Diseases of the Brain. Dementia is one of the classical Mitochondrial diseases of the brain, which is characterized by behavioural, motor and cognitive impairments. Altered mitochondrial function has been implicated in the pathogenesis of various neurodegenerative diseases such as Parkinson's disease, Alzheimer's disease, Friedrich's Ataxia, and Amylotrophic Lateral sclerosis (73). More recently it has been shown that autism and Chronic fatigue syndrome also represent mitochondrial diseases of the brain.
Decreased a-ketoglutarate activity in the brain in Alzeimer's. Further reduction in energy output in the brain of people with AD is seen in the reduced level of a-ketoglutarate in the brain of people with AD potentially implicating reduced levels of vitamin B1, vitamin B2 and lipoate (74, 75).
Beta Amyloid Plaques. Whilst it is not possible to see these directly, inspection of the brains of AD reveals the presence of "beta-amyloid" plaques. These are postulated to be caused by improper processing of "amyloid precursor protein to yield beta amyloid".
Neurofibrillary Tangles. Microscopic inspection of the brain of AD patients reveals the presence of "neurofibrillary tangles". These have been postulated to occur due to improper functioning of a protein called "Tau" which is responsible for stabilizing microtubules within the nerve cells. This improper processing then causes disruption of the microtubule structures leading to neurofibrillary tangles.
An excellent video describing this process can be found at alzheimers-disease-video.
The potential reasons for, or causes of, the deficiencies outlined above will be discussed on the relevant web-page.
References
-
Sott etal, 2005. Randomized controlled trial of homocysteine-lowering vitamin treatment in elderly patients with vascular disease PMID: 16332666
-
Seshadri etal, 2002.Plasma homocysteine as a risk factor for dementia and Alzheimer's disease. PMID: 11844848
-
Zeng etal, 2015 Homocysteine, endothelin-1 and nitric oxide in patients with hypertensive disorders complicating pregnancy. PMID: 26923880
-
Kamat etal, 2015 Homocysteine induced cerebrovascular dysfunction: Al link to Alzheimer's disease etiology. PMID: 26157520
-
Wang etal, 2014 Meta-analysis of plasma homocysteine content and cognitive function in elderly patients with Alzheimer's disease. PMID: 25664013
-
Madsen etal, 2015 Higher homocysteine associated with thinner cortical gray matter in 803 participants from the Alzheimer's disease neuroimaging initiative. PMID: 25444607
-
Choi etal, 2014 Association of homocysteine with hippocampal volume independent of cerebral amyloid and vascular burden. PMID: 25424964
-
de Jager 2014 Critical levels of brain atrophy associated with homocysteine and cognitive decline. PMID:24927906
-
Pushpakumar etal, 2014 Endothelial dysfunction: the link between homocysteine and hydrogen sulfide. PMID: 25008153
-
Ravaglia etal, 2004 Homocysteine and cognitive performance in healthy elderly subjects. PMID: 15207433
-
Ravaglia etal, 2003 Homocysteine and cognitive performance in healthy elderly community dwellers in Italy. PMID: 12600859
-
Adunsky etal, 2005 Plasma homocysteine levels and cognitive status in long-term stay geriatric patients.... PMID: 15680497
-
Koike etal, 2008 Raised homocysteine and low folate and vitamin B12 concentrations predict cognitive decline in community-dwelling older Japanese adults. PMID: 18835068
-
Li etal, 2008 Cognitive performance and plasma levels of homocysteine, vitamin B12, folate and lipids in patients with Alzheimer disease. PMID: 18931498
-
Issac etal, 2015 Vitamin B12 deficiency: an important reversible co-morbidity in meuropsychiatric manifestations. PMID: 25722058
-
Morris etal, 2012. Vitamin B-12 and folate status in relation to decline in scores on the mini mental state examination in the Framingham heart study PMID: 22788704
-
Inada etal 1982 Cobalamin contents of the brains in some clinical and pathological states. PMID: 7160970
-
Zhang etal, 2016 Decreased levels of vitamin B12 in aging, autism and schizophrenia. PMID: 26799654
-
Ikeda etal 1992 Treatment of Alzheimer-type dementia with intravenous mecobalamin. PMID: 1638584
-
Eastley etal, 2000 Vitamin B12 deficiency in dementia and cognitive impairment: the effects of treatment on neuropsychological function. PMID: 10713580
-
Manassiev 2014 Re:vitamin B12 deficiency www.bmj.com/content/349/bmj.g5226rr/763712
-
Fisar etal, 2016 Mitochondrial respiration in the platelets of patients with Alzheimer's disease PMID 2671932
-
Mattis J, Sehgal A. Circadian Rhythms, Sleep, and Disorders of Aging. Trends Endocrinol Metab. 2016 Apr;27(4):192-203. doi: 10.1016/j.tem.2016.02.003. Epub 2016 Mar 3. PMID: 26947521; PMCID: PMC4808513.
-
Holingue C, Wennberg A, Berger S, Polotsky VY, Spira AP. Disturbed sleep and diabetes: A potential nexus of dementia risk. Metabolism. 2018 Jul;84:85-93. doi: 10.1016/j.metabol.2018.01.021. Epub 2018 Feb 1. PMID: 29409842; PMCID: PMC5995651.
-
Blackman J, Swirski M, Clynes J, Harding S, Leng Y, Coulthard E. Pharmacological and non-pharmacological interventions to enhance sleep in mild cognitive impairment and mild Alzheimer's disease: A systematic review. J Sleep Res. 2021 Aug;30(4):e13229. doi: 10.1111/jsr.13229. Epub 2020 Dec 2. PMID: 33289311; PMCID: PMC8365694.
-
Porter VR, Buxton WG, Avidan AY. Sleep, Cognition and Dementia. Curr Psychiatry Rep. 2015 Dec;17(12):97. doi: 10.1007/s11920-015-0631-8. PMID: 26478197.
-
Chen JC, Espeland MA, Brunner RL, Lovato LC, Wallace RB, Leng X, Phillips LS, Robinson JG, Kotchen JM, Johnson KC, Manson JE, Stefanick ML, Sarto GE, Mysiw WJ. Sleep duration, cognitive decline, and dementia risk in older women. Alzheimers Dement. 2016 Jan;12(1):21-33. doi: 10.1016/j.jalz.2015.03.004. Epub 2015 Jun 15. PMID: 26086180; PMCID: PMC4679723.
-
Henry A, Katsoulis M, Masi S, Fatemifar G, Denaxas S, Acosta D, Garfield V, Dale CE. The relationship between sleep duration, cognition and dementia: a Mendelian randomization study. Int J Epidemiol. 2019 Jun 1;48(3):849-860. doi: 10.1093/ije/dyz071. PMID: 31062029; PMCID: PMC6659373.
-
Sabia S, Fayosse A, Dumurgier J, van Hees VT, Paquet C, Sommerlad A, Kivimäki M, Dugravot A, Singh-Manoux A. Association of sleep duration in middle and old age with incidence of dementia. Nat Commun. 2021 Apr 20;12(1):2289. doi: 10.1038/s41467-021-22354-2. PMID: 33879784; PMCID: PMC8058039.
-
Xu J, Wang LL, Dammer EB, Li CB, Xu G, Chen SD, Wang G. Melatonin for sleep disorders and cognition in dementia: a meta-analysis of randomized controlled trials. Am J Alzheimers Dis Other Demen. 2015 Aug;30(5):439-47. doi: 10.1177/1533317514568005. PMID: 25614508.
-
Peter-Derex L, Yammine P, Bastuji H, Croisile B. Sleep and Alzheimer's disease. Sleep Med Rev. 2015 Feb;19:29-38. doi: 10.1016/j.smrv.2014.03.007. Epub 2014 Apr 3. PMID: 24846773.
-
Sumsuzzman DM, Choi J, Jin Y, Hong Y. Neurocognitive effects of melatonin treatment in healthy adults and individuals with Alzheimer's disease and insomnia: A systematic review and meta-analysis of randomized controlled trials. Neurosci Biobehav Rev. 2021 Aug;127:459-473. doi: 10.1016/j.neubiorev.2021.04.034. Epub 2021 May 3. PMID: 33957167.
-
Tseng PT, Zeng BY, Chen YW, Yang CP, Su KP, Chen TY, Wu YC, Tu YK, Lin PY, Carvalho AF, Stubbs B, Matsuoka YJ, Li DJ, Liang CS, Hsu CW, Sun CK, Cheng YS, Yeh PY, Shiue YL. The Dose and Duration-dependent Association between Melatonin Treatment and Overall Cognition in Alzheimer's Dementia: A Network Meta- Analysis of Randomized Placebo-Controlled Trials. Curr Neuropharmacol. 2022;20(10):1816-1833. doi: 10.2174/1570159X20666220420122322. PMID: 35450525; PMCID: PMC9886806.
-
Potes Y, Cachán-Vega C, Antuńa E, García-González C, Menéndez-Coto N, Boga JA, Gutiérrez-Rodríguez J, Bermúdez M, Sierra V, Vega-Naredo I, Coto-Montes A, Caballero B. Benefits of the Neurogenic Potential of Melatonin for Treating Neurological and Neuropsychiatric Disorders. Int J Mol Sci. 2023 Mar 2;24(5):4803. doi: 10.3390/ijms24054803. PMID: 36902233; PMCID: PMC10002978.
-
Yang etal, 2016 Neuroprotection of Coenzym Q10 in neurodegenerative diseases PMID 26311425
-
Momiyama, 2014 Serum coenzyme Q10 levels as a predictor of dementia in a Japanese general population PMID 25463069
-
Yamagishi etal, 2014 Serum coenzyme Q10 and risk of disabling dementia the circulatory risk in communities PMID 25463064
-
Isobe etal, 2009 Increase in the oxidized/total coenzyme Q-10 ratio in the cerebrospinal fluid of Alzheimer's disease patients PMID 19907182
-
Chapman etal, 2016 Association between metformin and vitamin B12 deficiency in patients with type 2 diabetes... PMID 27130885
-
Harper et al, 2016 Brain levels of high-energy phosphate metabolites and executive function in geriatric depression. PMID: 26891040
-
Parks et al, 2013 Teaching neuroimages: neurodegeneration with brain iron accumulation in aceruloplasminemia. PMID: 24218322
-
Crespo etal, 2014 Genetic and biochemical markers in patients with Alzheimer's disease support a concerted systemic iron homeostasis dysregulation. PMID: 24199959
-
Guerreiro etal, 2015 Decrease in APP and Cp mRNA expression supports impairment of iron export in Alzheimer's disease patients. PMIT: 26209012
-
Milward etal, 2010 A cross-sectional community study of serum iron measures and cognitive status in older adults. PMID: 20182059
-
Roschel H, Gualano B, Ostojic SM, Rawson ES. Creatine Supplementation and Brain Health. Nutrients. 2021 Feb 10;13(2):586. doi: 10.3390/nu13020586. PMID: 33578876; PMCID: PMC7916590.
-
Ozaki T, Yoshino Y, Tachibana A, Shimizu H, Mori T, Nakayama T, Mawatari K, Numata S, Iga JI, Takahashi A, Ohmori T, Ueno SI. Metabolomic alterations in the blood plasma of older adults with mild cognitive impairment and Alzheimer's disease (from the Nakayama Study). Sci Rep. 2022 Sep 8;12(1):15205. doi: 10.1038/s41598-022-19670-y. PMID: 36075959; PMCID: PMC9458733.
-
Snow WM, Cadonic C, Cortes-Perez C, Adlimoghaddam A, Roy Chowdhury SK, Thomson E, Anozie A, Bernstein MJ, Gough K, Fernyhough P, Suh M, Albensi BC. Sex-Specific Effects of Chronic Creatine Supplementation on Hippocampal-Mediated Spatial Cognition in the 3xTg Mouse Model of Alzheimer's Disease. Nutrients. 2020 Nov 23;12(11):3589. doi: 10.3390/nu12113589. PMID: 33238473; PMCID: PMC7700653.
-
Mihara M, Hattori N, Abe K, Sakoda S, Sawada T. Magnetic resonance spectroscopic study of Alzheimer's disease and frontotemporal dementia/Pick complex. Neuroreport. 2006 Mar 20;17(4):413-6. doi: 10.1097/01.wnr.0000203353.52622.05. PMID: 16514368.
-
Raven etal, 2013 Increased iron levels and decreased tissue integrity in hippocampus of Alzheimer's disease detected in vivo with magnetic resonance imaging. PMID: 23792695
-
Yavuz etal, 2012 Iron deficiency can cause cognitive impairment in geriatric patients. PMID: 22546776
-
Liu etal, 2011 Iron promotes the toxicity of amyloid beta peptide by impeding its ordered aggregation. PMID: 21147772
-
Lee etal, 2009 A disruption in iron-sulfur centre biogenesis.... PMIT: 19290777
-
Ayton etal, 2015 Ferritin levels in the cerebrospinal fluid predict Alzheimer's disease outcomes and are regulated by APOE. PMID: 25988319
-
Armin etal 2010 In utero iron status and auditory nural maturation in premature infants as evaluated by auditory brainstem response PMID 19939407
-
Kwik-Uribe etal, 2000 Chronic marginal iron intakes during early development in mice result in persistent changes in dopamine metabolism and myelin composition.PMID 11053527
-
Choudhury etal 2015 Latent iron deficiency at birth influences auditory neural maturation in late preterm and term infants PMID 26310540
-
Armony-Sivan etal 2004 Iron status and neurobehavioral development of premature infants PMID 15318248
-
Wenger etal, 2017 Effect of iron deficiency on simultaneous measures of behavior, brain activity and energy.. PMID 28784049
-
Zumbrennen-Bullough etal 2014 Abnormal brain iron metabolism in Irp2 deficient mice is associated with mild neurological and behavioral impairments PMID 24896637
-
Georgieff 2008 The role of iron in neurodevelopment: fetal iron deficiency and the developing hippocampus PMID PMC2711433
-
Porter etal, 1994 The use of serum ferritin estimation in the investigation of anaemia in patients with rheumatoid arthritis Clin Exp Rheumatol. 1994 Mar-Apr;12(2):179-82
-
Finch etal, 1976 Iron deficiency in the rat. Physiological and biochemical studies of muscle dysfunction. PMC: 333200
-
Mangialasche etal, 2015 Lymphocytic mitochondrial aconitase activity is reduced in Alzheimer's disease and mild cognitive impairment. PMID: 25322927
-
Raukas etal, 2012 Mitochondrial oxidative stress... PMID: 22962855
-
Yarian etal 2006 Aconitase is the main functional target of aging in the citric acid cycle of kidney mitochondria in mice. PMID: 16289253
-
Corrigan etal, 1998 Abnormal content of N-6 and N-3 long-chain unsaturated fatty acids in the phosphoglycerides and cholesterol esters of parahippocampal cortex from Alzheimer's disease patients and its relationship to acetyl CoA content. PMID: 9698673
-
Kish etal, 1992 Brain cytochrome oxidase in Alzheimer's disease. PMID: 1321237
-
He etal, 2014 Hydrogen sulfide improves spatial memory impairment and decreases production of AB.... PMID: 24412510
-
Sonntag,etal, 2017 Late-onset Alzheimer's disease is associated with inherent changes in bioenergetics profiles. NATR s41587
-
Chen etal, 2015 Neuroprotective effects of hydrogen sulfide and the underlying signaling pathways. PMID: 25528761
-
Kamat etal, 2015 Hydrogen sulfide ameliorates homocysteine-induced Alzheimer's disease-like pathology..... PMID 26019015
-
Wang etal, 2014 Role of hydrogen sulfide in secondary neuronal injury. PMID: 24239876
-
Sen etal 2010 Homocysteine to hydrogen sulfide or hypertension. PMID: 20387006
-
Selley 2003 Increased concentrations of homocysteine and asymmetric dimethylarginine and decreased concentrations of nitric oxide in the plasma of patients with Alzheimer's disease. PMID: 12928408
-
Zeng etal, 2015 Homocysteine, endothelin-1 and nitric oxide in patients with hypertensive disorders complicating pregnancy. PMIT: 26823880
-
Lane etal, 2006 Targeting acetylcholinesterase and butylcholinesterase in dementia. PMID: 16083515
-
Tajeddinn etal, 2016 Association of platelet serotonin levels in Alzheimer's disease with clinical and cerebrospinal fluid markers. PMID: 27163811
-
Vermeiren etal, 2015 The monoaminergic footprint of depression and psychosis in dementia with Lewy bodies compared to Alzheimer's disease. PMID: 24717350
-
Werner etal, 2016 Serotonergic drugs: Agonists/antagonists at specific serotonergic subreceptors for the treatment of cognitive, depressant and psychotic symptoms in Alzheimer's disease. PMID: 26818863
-
Puranen etal, 2016 Incidence of antidepressant use in community-dwelling persons with and without Alzheimer's disease...PMID: 26924266
-
Cardoso etal, 2014 Selenium status in elderly: relation to cognitive decline PMID 25220532
-
Akbaraly etal, 2007 Plasm Selenium over time and cognitive decline in the elderly Epidemiology, Lippinscott. 18, 52-58
-
Haratake etal, 2013 Elevated amyloid-B plaque deposition in dietary selenium-deficient Tg2576 transgenic mice PMID 23456036
-
Nasrabady etal, 2018 White matter changes in Alzheimer's disease: a focus on myelin and oligodendrocytes PMC 5834839
-
Gáll Z, Székely O. Role of Vitamin D in Cognitive Dysfunction: New Molecular Concepts and Discrepancies between Animal and Human Findings. Nutrients. 2021 Oct 20;13(11):3672. doi: 10.3390/nu13113672. PMID: 34835929; PMCID: PMC8620681.
-
Morello M, Landel V, Lacassagne E, Baranger K, Annweiler C, Féron F, Millet P. Vitamin D Improves Neurogenesis and Cognition in a Mouse Model of Alzheimer's Disease. Mol Neurobiol. 2018 Aug;55(8):6463-6479. doi: 10.1007/s12035-017-0839-1. Epub 2018 Jan 9. PMID: 29318446; PMCID: PMC6061182.
-
Landel V, Annweiler C, Millet P, Morello M, Féron F. Vitamin D, Cognition and Alzheimer's Disease: The Therapeutic Benefit is in the D-Tails. J Alzheimers Dis. 2016 May 11;53(2):419-44. doi: 10.3233/JAD-150943. PMID: 27176073; PMCID: PMC4969697.
-
Szczechowiak K, Diniz BS, Leszek J. Diet and Alzheimer's dementia - Nutritional approach to modulate inflammation. Pharmacol Biochem Behav. 2019 Sep;184:172743. doi: 10.1016/j.pbb.2019.172743. Epub 2019 Jul 26. PMID: 31356838.
-
Yadav et al, 2019 Protein misfolding diseases and therapeutic approaches. Curr Protein Pept Sci, 20P1226-1245
-
Gamez et al, 2018 Protein misfolding diseases: Prospects of Pharmacological treatment. Clin Genet, 93; 450-458
-
Wankhede et al, 2022. Involvement of molecular chaperone in protein mis-folding brain diseases. 147l 116247
-
Pan, Y., Wallace, T. C., Karosas, T., Bennett, D. A., Agarwal, P., & Chung, M. (2024). Association of egg intake with Alzheimer’s dementia risk in older adults: The rush memory and aging project. The Journal of nutrition, 154(7), 2236-2243.
-
Oikonomidi, A., Lewczuk, P., Kornhuber, J., Smulders, Y., Linnebank, M., Semmler, A. and Popp, J. (2016), Homocysteine metabolism is associated with cerebrospinal fluid levels of soluble amyloid precursor protein and amyloid beta. J. Neurochem., 139: 324-332. https://doi.org/10.1111/jnc.13766
-
Soros etal, 2013 Antihypertensive treatment can prevent stroke and cognitive decline PMID 23247612
-
Ahmed etal 2019 Vitamin B12 deficiency and hyperhomocysteinaemia in outpatients with stroke or transient ischaemic attack. PMID 30670531
-
Liquori etal 2016 Cerebrospinal fluid lactate levels and brain [18f]FDG PET hypometabolism with the default mode network in Alzhemeir's Diseas. PMID 27221635
-
Chen and Zhong 2013 Decoding Alzheimer's disease from perturbed cerebral glucose metabolism: Implications for diagnostic and therapeutic strategies. PMID 23850509
-
Chaturvedi and Beal 2013 Mitochondrial diseases of the brain. PMID 23567191
-
Furst et al 2012 Cognition, glucose metabolism and amyloid burden in Alzheimer's disease. Neurobiol aging 33, 215-225
-
Banerjee et al 2016 Mild mitochondrial deficits....pathobiology of Alzheimer's disease. PMID 26923918
Copyright © 2018 B12 Oils. All Rights Reserved.
Reproduction in whole or in part in any form or medium without express written
permission is prohibited